




Por Marco Gonzalez

Lamitos da Formação Sousa, do Cretáceo inferior, encontrados no Vale dos Dinossauros, em Sousa, Pernambuco, Brasil.
Esta formação apresenta gretas de contração de paleoclima semiárido com alternância de estações chuvosas e secas e é marcada por pegadas de Iguanodonte [Medeiros, 2008].
(Crédito da imagem: Rocmayer).
Uma história climática da Terra no Mesozoico dá uma visão geral do clima e de suas interações com a geologia e a biologia entre ~252 milhões e ~66 milhões de anos atrás. Esta etapa da existência do nosso planeta durou ~186 milhões de anos, constituindo-se em uma era com clima quente e dominada por animais de grande porte, onde os continentes com seus relevos aplainados foram se aproximando dos contornos atuais.
4.1. Introdução
O Mesozoico, antes chamado de "Secundário" e conhecido como "Era dos Dinossauros", teve seu nome proposto em 1841 pelo geólogo britânico John Phillips (1800-1874), adotando a junção grega do adjetivo "mesos" ("meio") com o substantivo "zoe" ("vida"). [Rudwick, 2008; etymonline, 2018; Morrel, 2001]. É subdividido nos seguinte períodos:
O Mesozoico, antes chamado de "Secundário" e conhecido como "Era dos Dinossauros", teve seu nome proposto em 1841 pelo geólogo britânico John Phillips (1800-1874), adotando a junção grega do adjetivo "mesos" ("meio") com o substantivo "zoe" ("vida"). [Rudwick, 2008; etymonline, 2018; Morrel, 2001]. É subdividido nos seguinte períodos:
- Triássico, período com clima árido influenciado pela proximidade do Pangeia à linha do equador, é marcado pela evolução inicial dos mamíferos. Teve seu nome cunhado em 1834, pelo geólogo alemão Friedrich August von Alberti (1795-1878). Ele fez referência ao substantivo grego "trias" ("tríade, o número três") para caracterizar a subdivisão de três estratos típicos encontrados na Alemanha: Red Beds, gredas e folhelhos negros [Alberti, 1834; Speer et al, 1997; etymoline, 2024; NPS, 2025]
- Jurássico, período com clima quente que viu evoluir os dinossauros e as primeiras aves. Seu nome foi usado pela primeira vez em 1795 pelo geógrafo, polimata, naturalista, explorador e filósofo alemão Friedrich Wilhelm Heinrich Alexander von Humboldt (1769-1859), o Barão de Humboldt, para distinguir estratos das Montanhas Jura do norte da Suíça [Waggoner et al, 1995; NPS, 2023].
- Cretáceo, período típico de clima de estufa, é marcado pelo fim do domínio dos dinossauros na Terra. Seu nome foi proposto em 1882 pelo estadista e geólogo belga Jean Baptiste Julien d'Omalius d'Halloy (1783-1875) e tem origem no latim "creta" ("greda, giz") com inspiração nas grandes quantidades de rocha calcária deste período na Europa Ocidental [Wang, 2021; NPS, 2023; Osterloff, 2024]

Tabela cronoestratigráfica do Mesozoico.
(Baseada em: ICS)
O Mesozoico presenciou transformações fundamentais na evolução climática da Terra em direção ao seu estado atual e foi marcado pela fragmentação do Pangeia e por inovações bióticas significativas [Landwehrs et al, 2021]

Eventos climáticos, geológicos e biológicos do Mesozoico.
Evolução da vida a...i: descrições na tabela que pode ser visualizada a seguir.
Extinções em massa (⁂) do Permiano-Triássico [Preto et al, 2010; Wilson et al, 2019], do final do Triássico [Hautmann, 2012] e do Cretáceo-Paleógeno [Archibald, 2002; Chiarenza et al, 2020; Scotese et al, 2024].
Curva clássica (Famílias) da diversidade de Sepkoski de famílias de invertebrados marinhos [Servais et al, 2009; Sepkoski Jr., 2016].
Aves (Aves) [Chen et al, 2024], Mamíferos (Mam) [Cabreira et al, 2022a], Dinossauros (Dino) [Nesbitt et al, 2013] e Répteis (Rep) [Keighley et al, 2008; Strauss, 2019].
Deriva Continental: Pangeia (P), Gondwana (G), Laurásia (L) e insipientes América do Norte (1), Europa (2), América do Sul (3), África (4), Antártica (5) e Austrália (6) [Algol, 2016; Scotese, 2016; Doppel, 2024].
Nível do mar global [Murphy e Nance, 2013]
Principais Grandes Províncias Ígneas (LIPs): Traps Siberianos e Emeishan (1), Wrangellia (2), Atlântico Central (3), Karoo-Ferrar (4), Mikabu (5), Parana-Etendeka (6), Alto Ártico (7), Caribe e Madagascar (8) e Atlântico Norte e Traps de Deccan (9) [Tominaga e Hara, 2021; Boscaini et al, 2022; Percival et al, 2024; Nechaev et al, 2022].
Curvas da luminosidade do Sol [Ramstein et al, 2019], do ciclo dia-noite [Arbab, 2003], da temperatura [Judd e al, 2024] e das concentrações de O₂ [Royer, 2014] e CO₂ [Royer, 2014] atmosféricos abstraindo incertezas, pretendendo dar uma ideia aproximada das variações dos valores médios globais.
Tabela da evolução da vida no Mesozoico
A árvore filogenética
simplificada dos principais tetrápodes
do Mesozoico

4.2. Uma história climática do Mesozoico
4.2.1. Triássico
| De 251,9 milhões a 201,4 milhões de anos atrás | |
| Luminosidade do Sol | de 98 a 98,3% da atual |
| Duração do ciclo dia-noite | de 22,5 a 22,8 horas |
| Temperatura | de 21 a 23°C com mínima de 19,2 e máxima de 28,9°C |
| Concentração de O₂ na atmosfera | de 23 a 19,5% com mínima de 18,7% |
| Quantidade de CO₂ na atmosfera | de 750 a 500 ppm |
4.2.1.1. Clima não zonal e efeito estufa no início do Triássico
A extinção em massa Permiano-Triássico, a formação e a erosão das Traps Siberianas
e outros fatores se combinaram para preservar um clima severo prolongado no início do Triássico [Wilson et al, 2019]. Em geral, havia oscilações climáticas contidas numa estrutura de padrão não zonal,
com forte sistema global de monções causando efeitos evidentes no contexto do paleoceano Tétis,
relacionado à agregação do Pangeia [Preto et al, 2010].
A produção de carbonato se manteve elevada no início do Triássico, pelo menos na porção ocidental do Tétis, ainda que os metazoários
construtores de recifes tenham sido talvez o grupo mais afetado pela extinção em massa do início do período. O clima de estufa do Triássico inferior teria provocado empobrecimento da fauna e da flora, embora tenha havido notáveis evoluções [Preto et al, 2010]. No início do Triássico as coníferas
se expandiram, mas, onde o clima se tornou mais seco, as florestas deram lugar a vastas pradarias de samambaias [Davis, 2024].
4.2.1.2. Recuperações da extinção do Permiano-Triássico
Nestas recuperações, mesmo com aumento da competição por recursos, houve expansão da biodiversidade. Houve atrasos no início do Triássico Inferior, talvez consequência da persistência de condições ambientais difíceis, ao contrário da rápida expansão na fase final do Triássico inferior [Erwin, 2007]. Porém, o que surgiu na biosfera não foi tão diferente do que existia antes [Davis, 2024].
No Triássico inferior, o vertebrado mais comum passou a ser o pequeno e herbívoro sinapsídeo,
ou réptil semelhante a mamífero, chamado Lystrosaurus [Davis, 2024].

Reconstrução do Lystrosaurus georgi.
É possível que os Lystrosaurus tenham se adaptado à vida subterrânea em túneis escavados por eles para escapar da atmosfera cheia de poeira e cinzas do Triássico [Newitz, 2013].
(Crédito da imagem: Dimitri Bogdanov).
Entre 250 milhões e 246 milhões de anos atrás, os primeiros ictiossauros
foram para os mares, constituindo um grupo que eventualmente dominaria os oceanos [Davis, 2024]. Um ictiossauro notável pelo tamanho é o Shonisaurus [Green, 2012].
Nesta recuperação, uma das principais transições da fauna na Terra ocorreu com os répteis arcossauromorfos, insignificantes no ecossistema até se tornarem vertebrados dominantes [Ezcurra e Butler, 2018].
Eles floresceram e se diversificaram à medida que preenchiam nichos ecológicos vazios. Alguns deles prenunciavam características de futuros dinossauros, como teto de crânio espesso [Benton, 2016].
Trazendo consequências para as faunas do Mesozoico e modernas, os ecossistemas globais foram transformados, em várias fases, pela seguinte sequência de diversificações [Ezcurra e Butler, 2018]:
- constituição, como ponto de partida, de uma fauna de desastre pós-extinção;
- diversificação significativa, mas mal entendida e amostrada, com taxas evolutivas bem elevadas; e
- aumento acentuado na contagem de espécies, com abundância e disparidade simultâneas à estabilização do ecossistema global ~5 milhões de anos após a extinção.
Quanto às plantas, seus fósseis desde final do Permiano revelam recuperações efetivas até o final do Triássico médio. O clima foi essencial contribuindo com baixo gradiente de temperatura da linha do equador para os polos. Um dos primeiros sinais de recuperação na flora, constatados no norte da China, foi a proliferação da Pleuromeia,
planta que foi a característica mais distintiva do Triássico Inferior em toda a Eurásia [Grauvogel-Stamm et al, 2005].

Reconstrução da Pleuromeia sternbergii
do Triássico da Alemanha.
Nos continentes do sul há indícios de que a Pleuromeia tenha sido uma planta pioneira oportunista que aproveitou a vacuidade do ambiente pós-crise para proliferar, preparando locais para recolonização ao sobreviver em refúgios [Grauvogel-Stamm et al, 2005].
(Crédito da imagem: Duijnstee e Bonner).
A recuperação da flora prosseguiu no início do Triássico médio com o ressurgimento das coníferas e o retorno das cicadáceas
e das pteridospermas,
com subsequente progressiva modernização evolutiva [Grauvogel-Stamm et al, 2005].
4.2.1.3. O Evento Pluvial Carniano
O clima do Triássico em geral pode ser considerado relativamente quente e seco, com exceção do intervalo quente mas chuvoso denominado Evento Pluvial Carniano (EPC)
[Corecco et al, 2024] que aconteceu há ~233 milhões de anos [Lu et al, 2021].

Durante o EPC, as bacias do Tétis ocidental foram inundadas, sendo depositados os folhelhos da Formação Reingraben com esporos fossilizados
[Corso et al, 2015]. O EPC provocou aumento da umidificação e da fossilização destes folhelhos. O clima induziu a baixa oxigenação dos sedimentos de fundo com consequente laminação [Lukeneder e Lukeneder, 2023].
(Crédito da imagem: Lukeneder e Lukeneder).
O EPC ocorreu durante uma fase de "mares de aragonita",
com a geoquímica da água do mar semelhante à atual. Entre suas causas são consideradas as seguintes [Preto et al, 2010]:
- alterações na dinâmica atmosférica e na circulação oceânica impulsionadas pela tectônica de placas;
- intensificação da monção global devido à agregação máxima de continentes; e
- erupção vulcânica com formação de grande província ígnea.
Esta última causa associa o EPC ao vulcanismo de larga escala da Província Ígnea de Wrangellia.
Acredita-se que este período pluvial foi desencadeado pela emissão volumosa de CO₂ vulcânico, com consequente aquecimento global e aumento de clima de mega-monção [Lu et al, 2021]. O vulcanismo de Wrangellia coincidiu com o desenvolvimento de condições redutoras nas águas profundas do paleoceano Pantalassa
[Tomimatsu et al, 2023]. Posteriormente, as condições climáticas parecem ter se estabilizado com pequenas alterações [Preto et al, 2010].
4.2.1.4. Crocodilianos
No final do Triássico Médio, com o clima cada vez mais quente e seco, os sinapsídeos estavam em declínio e uma gama diversificada de arcossauros havia surgido.
Os arcossauros faziam parte da linhagem dos diapsídeos,
incluindo grupos do Mesozoico bem-sucedidos, como os crocodilianos
e as tartarugas [Davis, 2024].
Na linhagem dos crocodilianos, no Triássico, destacaram-se o rincossauro
e o fitossauro [Renton e Repine, 2016].
Os primeiros ancestrais dos crocodilianos nem sempre foram predadores aquáticos de emboscada. Há 220 milhões de anos a maioria dos seus parentes corria em terra firme. A linha de crocodilianos incluía uma grande variedade de formas, desde herbívoros semelhantes a tanques blindados até grandes predadores. Um arcossauro da linha dos crocodilianos do Triássico superior era o Poposaurus gracilis, que chegou a ser confundido com dinossauro [Irmis, 2019].

Restauração do Poposaurus gracilis.
Sua "caminhada em altura" em baixa velocidade era viabilizada pelos membros posteriores oscilando em plano mais próximo do centro de massa durante todo o ciclo de cada passo. A postura mais ereta foi permitida por um conjunto de mudanças anatômicas em pés, pernas e pélvis, além das que fortaleceram o tronco e melhoram a flexão da coluna vertebral [Gauthier et al, 2011]. Esta postura afasta a imagem de répteis submersos à espreita nos pântanos e rios quentes e úmidos do período [Irmis, 2019]
(Crédito da imagem: Smokeybjb).
4.2.1.5. Tartarugas
As verdadeiras tartarugas surgiram no Triássico superior e rapidamente alcançaram distribuição mundial e diversidade relativamente elevada [Szczygielski et al, 2025]. Dentre elas, há a Palaeochersis talampayensis da Argentina
e a Proganochelys quenstedti da Alemanha [Sterli et al, 2007].

Esta tartaruga vivia em região de sistemas deltaicos ou lacustres próximos à costa [Rieppel e Reisz, 1999]. Provavelmente adotou um habitat misto, já que a morfologia dos membros anteriores e os ossos da carapaça indicam que seria terrestre, enquanto a morfologia da carapaça e do fêmur sugerem que seria aquática [Lichtig e Lucas, 2021]
(Crédito da imagem: Claire Houck).
4.2.1.6. Dinossauros
Dentre os arcossauros bem-sucedidos do Mesozoico, incluem-se também os dinossauros e os pterossauros [Davis, 2024].
Os dinossauros evoluíram no único supercontinente que havia, o Pangeia, árido e inóspito cercado pelo oceano Pantalassa [NPS, 2023]. Estavam entre os sobreviventes da extinção do Permiano-Triássico e, com exceção do Evento Pluvial Carniano, viviam em condições climáticas únicas quentes e secas decorrentes da configuração continental. Nestas condições, os dinossauros começaram a se espalhar amplamente ocupando a maioria dos nichos da macrofauna de terra firme [Corecco et al, 2024].
O dinossauro mais antigo conhecido, Nyasasaurus parringtoni,
de 243 milhões de anos, resultou de uma combinação de características dos dinossauros em evolução, com a taxa de crescimento elevada dos dinossauros primitivos. Demonstrou que a fase inicial de aparecimento de novas espécies de dinossauros ocorreu em uma escala de tempo mais demorada do que se pensava, possivelmente iniciada 15 milhões de anos antes [Nesbitt et al, 2013].

Representação do Nyasasaurus indicando a posição de fragmentos esqueléticos parciais, um úmero e seis vértebras (em preto) do primeiro espécime encontrado e três vértebras cervicais (em azul) do segundo espécime.
(Crédito: KDS444)
Enquanto os arcossauros não dinossauros continuaram a dominar a maioria dos ambientes, pouco tempo depois de surgirem, os dinossauros teriam divergido em dois grupos principais: Saurischia
e
Ornithischia [Davis, 2024].
Os primeiros dinossauros primitivos eram pequenos animais bípedes, como o Coelophysis,
da mesma forma que seus ancestrais. Não eram parte importante da fauna que incluía anfíbios, répteis e animais semelhantes a mamíferos, onde se estacavam os blindados aetossauros [NPS, 2023].
No final do Triássico, porém, alguns dos primeiros dinossauros já eram impressionantes em tamanho. Alguns dos primeiros parentes dos saurópodes,
como Riojasaurus
e Lessemsaurus,
pesavam toneladas [Davis, 2024].
4.2.1.7. Pterossauros
Os Pterossauros foram os primeiros vertebrados a alçar voo. O mais antigo pterossauro é o Caelestiventus hanseni,
de ~208 milhões de anos [Britt et al, 2018].

Este pterossauro foi encontrado em sedimentos de um oásis de deserto no atual estado de Utah, EUA. Foi vítima de uma seca naquele oásis que aconteceu entre 210 milhões e 201 milhões de anos [Perkins, 2018].
(Crédito da imagem: Nobu Tamura).
No Triássico Superior, uma mudança no domínio entre os répteis semelhantes a mamíferos e os arcossauros tem várias teorias sobre sua causa, como estagnação evolutiva e competição em um clima que estava se tornando cada vez mais quente e seco. Há indícios de que os arcossauros foram mais capazes de preencher os nichos vazios deixados após a extinção de algumas das linhagens de sinapsídeos [Davis, 2024]. Eram tempos em que magmatismo e rifteamento preconizavam a fragmentação do Pangeia [de Lamotte et al, 2015].
4.2.1.8. Magmatismo e rifteamento passivo
O evento magmático relacionado às Traps Siberianas, iniciado no Permiano, avançou no Triássico até ~242 milhões de anos atrás. Neste período se desenvolveu o domínio do paleoceano Tétis que ficou vinculado a um episódio de rifteamento passivo [de Lamotte et al, 2015].
O início da fragmentação do Pangeia foi marcado, no Triássico, por este rifteamento passivo com consequente produção da Província Magmática do Atlântico Central (PMAC)
há ~200 milhões de anos. Estes eventos seriam associados, posteriormente, à abertura do Oceano Atlântico central [de Lamotte et al, 2015]. A PMAC é discutida adiante no contexto da extinção em massa do final do Triássico.

A Formação Blomidon, na Bacia de Fundy, compreende 200 a 300 m de arenito e argilito, intercalados ciclicamente, depositados em ambiente eólico e fluvial, em clima semiárido a árido [Tanner e Kyte, 2005]. Sobre estes sedimentos, o vulcanismo do PMAC parece ter ocorrido em período de tempo muito curto coerente com o desencadeamento da crise biótica na fronteira Triássico-Jurássico [Cirilli et al, 2009].
(Crédito da imagem: Michael C. Rygel).
4.2.1.9. Mamíferos primitivos
Com o clima quente e árido, no Triássico superior, os pequenos mamíferos primitivos, para evitar o calor do dia, tornaram-se exclusivamente noturnos (assim como atualmente a maioria deles). Provavelmente este hábito foi causado pela elevação da temperatura dos seus corpos, resultado da evolução da pelagem e do aumento da taxa de consumo de oxigênio por unidade de massa corporal [Lovegrove, 2019].
Esta alteração para sangue quente, a endotermia, no Triássico superior, há mais de 200 milhões de anos, indicou o início de um estilo de vida mais ativo. O sangue quente lhes disponibilizou novos comportamentos e habitats. Há indícios de que fizeram esta transição de forma abrupta, mas a origem da endotermia mamífera é uma questão não resolvida na paleontologia [Ashworth, 2022], já que a maioria das evidências fósseis é ambígua [Araújo et al, 2022]. Acredita-se que ancestrais de aves e de mamíferos tenham adquirido endotermia simultaneamente, envolvidos em uma corrida armamentista [Benton, 2021].
Muitos terapsídeos primitivos,
considerados ancestrais dos mamíferos, são amplamente usados em bioestratigrafia devido à sua abundância relativa e à distribuição global. Estes "répteis semelhantes a mamíferos" apresentam mistura de características reptilianas e mamíferas preservadas em seus fósseis. Foram os vertebrados terrestres dominantes durante grande parte do Paleozoico superior e do Mesozoico inferior e, portanto, dão testemunho da diversificação dos vertebrados em terra firme e no estabelecimento de ecossistemas terrestres modernos [Rubidge e Sidor, 2001].
O Brasilodon quadrangularis
é considerado o mamífero mais antigo conhecido no registro fóssil datado em 225,42 milhões de anos [Cabreira et al, 2022a].

Reconstrução do Brasilodon quadrangularis.
Foi encontrado na Sequência Candelária da Supersequência Santa Maria, no município de Faxinal do Soturno, Rio Grande do Sul, Brasil. Viveu em um clima úmido próximo a rios de baixa sinuosidade, deltas e lagos do sul da América do Sul [Cabreira et al, 2022a]. Provavelmente usava tocas como uma estratégia alternativa para se proteger dos dinossauros mais antigos conhecidos, já que era contemporâneo deles [Cabreira et al, 2022b].
(Crédito da imagem: Jorge Blanco).
Considerava-se o Eozostrodon, que habitava florestas da atual Europa ocidental,
como primeiro mamífero do registro fóssil. Porém, ele teria evoluído dos terapsídeos há 210 milhões de anos [Strauss, 2019] e portanto 15 milhões de anos depois do Brasilodon quadrangularis [Cabreira et al, 2022a]. De qualquer forma, como todos os mamíferos primitivos, estes provavelmente viviam no alto das árvores para não ser esmagados pelos grandes dinossauros [Strauss, 2019].
4.2.1.10. Extinção em massa do final do Triássico
A extinção em massa do final do Triássico (também chamada Triássico-Jurássico) parece estar associada à alteração climática relacionada à Província Magmática do Atlântico Central (PMAC) [Preto et al, 2010]. Esta extinção foi uma das cinco maiores na história da Terra [Hautmann, 2012], teve início há 201,5 milhões de anos e teria se estendido até a fronteira Triássico-Jurássico, durando entre 60 mil e 140 mil anos [Blackburn et al, 2013].
Suas causas, ainda não bem conhecidas [Davis, 2024], incluem as seguintes hipóteses [Hautmann, 2012]:
- impacto de corpo extraterrestre (com fracas evidências),
- alterações climáticas,
- mudanças do nível do mar,
- anoxia marinha,
- liberação de compostos tóxicos e
- acidificação da água do mar.
Com exceção da primeira, as demais podem ser incorporadas ao cenário de extinção vulcanogênica associado à atividade contemporânea da PMAC. Esta atividade anunciou a fragmentação do Pangeia após este supercontinente ter persistido por quase 150 milhões de anos. Informações sobre datações radiométricas indicam o início repentino das extrusões vulcânicas sincronizadas com as eliminações ocorridas na biosfera [Hautmann, 2012].

Localização da área da PMAC (em vermelho claro) no Pangeia.
Dados geocronológicos referentes ao derrame basáltico demonstram a correlação entre o vulcanismo e a extinção em massa. A liberação de magma e o fluxo atmosférico associado revelam quatro pulsos magmáticos ao longo de cerca de 600.000 anos, indicando vulcanismo expansivo mesmo enquanto a recuperação biológica estava em andamento [Blackburn et al, 2013].
(Imagem baseada em: Eltiamieli; Williamborg).
A atividade vulcânica iniciou há 201,8 milhões de anos e continuou com pulsos magmáticos. Aconteceu forte tectônica de rifte que acabou causando posteriormente a abertura do Atlântico central. O derrame basáltico se estendeu por uma área de mais de dez mil km² em ambos os lados da zona de rifte produzindo um volume de 2,3 milhões de km³ de material vulcânico e resultando em uma das maiores províncias ígneas da história da Terra [Hautmann, 2012].
Grande quantidade liberada de CO₂ e SO₂ teria provocado graves convulsões climáticas e contribuído principalmente para extinções marinhas [Davis, 2024]. Houve resfriamento de curto prazo inicial com aerossóis de SO₂ seguido de aquecimento global de longo prazo com efeito estufa com CO₂ [Hautmann, 2012].
No ambiente terrestre, há evidências de perturbação das comunidades vegetais, mas apenas algumas espécies desapareceram [Hautmann, 2012], como é o caso de vários grupos de plantas com semente [Vajda et al, 2023]. Outro caso de perda é o das turfeiras, tanto as costeiras quanto as próximas do mar epicontinental europeu e da margem europeia do Tétis. Nestes locais, os efeitos foram devastadores causados pela mudança climática com aumento da temperatura do ar e rápida elevação do nível do mar. Houve graves perdas principalmente de árvores e arbustos gimnospermas
adaptados a tais habitats. Esta devastação foi ainda amplificada por queda subsequente do nível do mar, viabilizando que oportunistas herbáceas
colonizassem um ambiente que estava estressado também pelo aumento da atividade de incêndios florestais e pela erosão intensificada do solo [Lindström, 2021].
Os vertebrados terrestres também sofreram [Hautmann, 2012]. Todos os arcossauros do Triássico foram extintos, exceto alguns pterossauros, crocodilianos e dinossauros [Davis, 2024]. Sobreviveram também os répteis semelhantes a mamíferos, os ancestrais dos anfíbios e os répteis marinhos [NPS, 2023]. Estas extinções abriram caminho para que os dinossauros sobreviventes assumissem nichos ecológicos desocupados e passassem a dominar a Terra [Chu, 2013].
No mar, foram eliminados os conodontes
e quase aniquilados corais e esponjas esfinctozoárias.
Ainda foram fortemente afetados braquiópodes,
bivalves,
gastrópodes
e foraminíferos.
Quase todos os grupos de amonóides foram extintos.
Os organismos construtores de recifes foram vítimas proeminentes [Hautmann, 2012].
Estima-se que a extinção do final do Triássico dizimou 76% das espécies marinhas e terrestres [Chu, 2013].
4.2.2. Jurássico
| De 201,4 milhões a 145 milhões de anos atrás | |
| Luminosidade do Sol | de 98,3 a 98,7% da atual |
| Duração do ciclo dia-noite | de 22,8 a 23,1 horas |
| Temperatura | de 23 a 27,2°C com mínima de 18°C |
| Concentração de O₂ na atmosfera | de 19,5 a 15,2% com mínima de 13,5% |
| Quantidade de CO₂ na atmosfera | de 500 a 540 ppm com mínima de 344 ppm e máxima em 714 ppm |
4.2.2.1. Deriva continental e vulcanismo
A abertura da porção central do Oceano Atlântico, com o início da ruptura do Pangeia no Jurássico inferior, foi acompanhada de vulcanismo generalizado acarretando alterações significativas na composição atmosférica, no clima e no ambiente do sistema de superfície da Terra [Zhou et al, 2022].
Algumas destas alterações são atribuídas à implantação da Grande Província Ígnea Karoo-Ferrar [Zhou et al, 2022],
cujos derrames basálticos são associados, além da ruptura do Gondwana, também à abertura do Oceano Índico [Jourdan et al, 2004]. Este eventos aconteceram devido a um rifteamento ativo
no Jurássico inferior desencadeado pela Karoo-Ferrar [de Lamotte et al, 2015]. O principal pulso de magnetismo de toda esta província apresenta sincronismo com pequeno episódio de extinção em massa no final do Jurássico inferior. Este sincronismo é revelado por datação de doleritos da Tasmânia [Ivanov et al, 2016].
(Crédito: Hannes Grobe).
Karoo-Ferrar sofreu influência das placas tectônicas do Pangeia devido à redução do movimento destas enquanto revertiam de direção. Simultaneamente ao desenvolvimento desta província ígnea, aconteceu o Evento Anóxico Oceânico Toarciano [Ruhl et al, 2022]
O vulcanismo induziu um aquecimento climático global que pode também ser correlacionado a condições climáticas mais úmidas, intemperismo continental intensificado, expansão de lagos e eutroficação.
No entanto, houve feedback negativo importante no ciclo global do carbono, aumentando o consumo de CO₂ em ambientes lacustres, resultando em restauração de climas e ambientes semelhantes aos que ocorriam antes do vulcanismo em algumas áreas [Zhou et al, 2022].
4.2.2.2. Recuperações da extinção em massa do final do Triássico
Houve grande renovação biótica nesta extinção [Allen et al, 2018], sendo a recuperação aparentemente mais rápida do que a que aconteceu na extinção do Permiano-Triássico. No início do Jurássico já havia ocorrido uma diversificação significativa entre invertebrados marinhos com ponto alto na porção intermediária do Jurássico inferior. Foi quando reapareceram os organismos recifais [Ros et al, 2011]. Houve recuperação relativamente lenta da fauna marinha na Europa atribuída à deficiência de oxigênio. Em contraste, houve recuperação quase instantânea de comunidades de águas profundas [Hautmann, 2012].
Nesta extinção, os amonóides quase desapareceram e somente no final do Jurássico médio a diversidade de suas conchas voltou a ser comparável à do Triássico superior. Estima-se que o tempo de recuperação total destes organismos tenha sido entre 30 milhões e 40 milhões de anos, sendo notável que um dos grupos mais bem-sucedidos de todos os tempos em termos de diversidade de espécies, longevidade e alcance global tenha demorado tanto para recuperar toda a sua gama de geometria de conchas [Smith et al, 2014]. Em compensação, os amonites
rapidamente se transformaram em invertebrados comuns no ambiente marinho, tornando-se fósseis índice
para estratos do Jurássico. Também os braquiópodes se recuperaram e prosperaram nos mares continentais quentes [Rafferty, 2011].

À esquerda, o fóssil de um amonite Asteroceras obtusum,
encontrado na Formação Lias, do Jurássico inferior, em Dorset, Inglaterra. À direita, a reconstrução de um amonite.
Asteroceras obtusum é amonite índice do intervalo chamado Zona Obtusum, que engloba rochas predominantemente marinhas do Jurássico inferior do nordeste da Europa [Edmunds et al, 2016]. Sedimentos da Zona Obtusum foram depositados em ambientes marinhos epicontinentais influenciados por flutuações do nível do mar e pelo clima quente e predominantemente úmido [Radley, 2008]. Nesta região, de forma mais ampla, há indícios de alternância de intervalos de clima quente úmido e semiárido [Munier et al, 2021].
Embora os bivalves tenham sido fortemente impactados nesta extinção, recuperaram-se durante o Jurássico Inferior embora não de forma homogênea. Moluscos bivalves carnívoros
(por exemplo, Septibranchia)
só apareceram no Jurássico médio [Ros et al, 2011].
Os arcossauromorfos, que eram altamente díspares e diversos no final do Triássico, sofreram grande impacto em sua evolução. Os arcossauros da linha dos crocodilianos diminuíram, enquanto os da linha das aves e mais notavelmente dos dinossauros e pterossauros passaram a dominar. Dentre os arcossauros, extinguiram-se os aetossauros, os fitossauros e os rauissúquios [Allen et al, 2018].
Acredita-se que o tamanho do corpo não esteja ligado à extinção dos arcossauromorfos. É o que mostram os dinossauros, que viriam a ser um dos grupos mais bem-sucedidos do Mesozoico exibindo uma ampla variedade de tamanhos corporais. Eles apresentaram variadas adaptações e ocuparam muitos nichos ecológicos diferentes [Allen et al, 2018].
4.2.2.3. Reestruturação das comunidades bentônicas no Jurássico inferior
A reestruturação abrangente das comunidades bentônicas,
um dos eventos paleoecológicos mais significativos do Mesozoico, é também conhecida como "Revolução Marinha do Mesozoico" [Lucas e Hunt, 2023]. É um evento que teve início no Jurássico (ou talvez mesmo no Triássico) com aumento na infaunalização
e na energia dos ecossistemas marinhos durante o Jurássico e o Cretáceo. Também cresceu a ação predatória durófaga
com resposta evolutiva das presas aumentando a robustez das conchas [Buatois et al, 2016; Lucas e Hunt, 2023].
A partir do Jurássico inferior, houve aparecimento relativamente rápido de novas espécies de equinoides em suas linhagens infaunais devido à colonização e à expansão em nichos desocupados, ou alternativamente, como uma resposta à crescente pressão predatória [Petsios et al, 2021].
O registro fóssil do Jurássico inferior revela uma resposta da comunidade bentônica ao aquecimento global e à anoxia marinha de longo prazo. Ocorreram dois principais eventos de renovação faunística coincidentes com o aumento tanto da temperatura quanto do nível do mar [Danise et al, 2013]:
- no início da anoxia, ocorreu extinção da maioria das espécies bentônicas e sobrevivência de algumas adaptadas para prosperar em condições de baixo oxigênio e,
- na recuperação, espécies recém evoluídas colonizaram os sedimentos moles reoxigenados evitando degradação ecológica.
4.2.2.4. Dinossauros
O início do Jurássico, com a abertura da porção norte do Oceano Atlântico e com a ocorrência de extensos derramamentos de lava [NPS, 2023], o clima era quente e seco e grande parte da terra firme era coberta por amplos desertos. Neste contexto, havia pouca diversidade entre animais e plantas [Reid-Collins e Brogan, 2025] e a chamada "Era dos Dinossauros" realmente foi um período de domínio destes animais em ecossistemas continentais. Eles se tornaram dominantes após a extinção do final do Triássico [Sues, 2016] com a remoção da maior parte dos competidores e com o início da sua diversificação [NPS, 2023]. Ou seja, a chamada "Era dos Dinossauros" realmente começou no início do Jurássico [Sues, 2016].
Entre os primeiros dinossauros do Jurássico inferior estão os heterodontossaurídeos,
que eram pequenos répteis que atingiam apenas ~2 metros de comprimento [Osterloff, 2025]. No Jurássico médio, quando o Pangeia se dividia em Laurentia ao norte e Gondwana ao sul, a precipitação aumentou devido à formação de mares entre as massas de terra, permitindo o crescimento de samambaias e florestas exuberantes. Foi o principal momento em que grupos de animais terrestres se diversificaram e se espalharam pela Terra, entre eles os dinossauros, surgindo os braquiossauros
e os megalossauros
[Reid-Collins e Brogan, 2025]. No Jurássico superior, são encontrados ossos e/ou pegadas de terópodes carnívoros,
enormes saurópodes e dinossauros blindados como os anquilossauros, com escudos,
os ornitópodes, herbívoros bípedes com bico,
e os estegossauros, com placas
[NPS, 2023].

Reconstrução do Stegosaurus stenops.
Encontrado nos sedimentos do Jurássico superior da Formação Morrison, Wyoming, EUA, este espécime parece ter sido bem preservado pelo soterramento rápido em um lago ou corpo d'água imediatamente após sua morte [Maidment et al, 2015]. A flora e os sedimentos da Formação Morrison (na porção onde o Stegosaurus stenops foi encontrado) indicam clima quente, sazonal e semiárido úmido [Parrish et al, 2004].
(Crédito da imagem: DBCLS).
No Jurássico Superior, algumas regiões propiciaram a preservação de um grande número de fósseis de dinossauros, como aquela representada pela Formação Morrison, na América do Norte. Ali são encontrados alossauros,
apatossauros,
brontossauros,
diplódocos
e estegossauros [NPS, 2023].

Reconstrução do Brontosaurus excelsus.
Este brontossauro foi encontrado na Formação Morrison, Colorado, EUA. Pegadas em sedimentos macios de rios e lagos indicam que, se podia viver em terra firme (estruturas de membros, coluna vertebral, mãos e pés eram adaptadas para suportar o grande peso do corpo), provavelmente também transitava por águas rasas (o que facilitaria suportar seu peso) [Osterloff, 2024].
(Crédito da imagem: UDL).
4.2.2.5. Outros répteis
A evolução dos pterossauros durante seus 149 milhões de anos de história (do Triássico superior ao final do Cretáceo) aconteceu por meio de adaptações envolvendo formato do crânio, dentição, tamanho do corpo e plano corporal (comprimento do pescoço, comprimento da cauda e envergadura das asas). As adaptações alimentares correlacionadas com a diversidade morfológica e filogenética ocorreram simultaneamente com a ascensão dos Pterodactiloides,
o grupo de pterossauros com maior diversidade de espécies do Jurássico superior ao final do Cretáceo [Zhou et al, 2017].
A evolução dos pterossauros foi aproximadamente paralela à dos dinossauros. Espécies pequenas e primitivas do final do Triássico gradualmente deram lugar a formas maiores e mais avançadas no Jurássico e no Cretáceo. As formas primitivas eram representadas pelos Ramforincoides.
Persistiram até o início e o meio do Jurássico e incluíam o eudimorfodonte (um dos primeiros pterossauros conhecidos),
o dorignato
e o ranforrinco [Strauss, 2019].

Restauração do pterossauro Dearc sgiathanach
e de seu ambiente no Jurássico médio da atual Escócia.
Foi encontrado no Membro Lonfearn da Formação Lealt Shale, na Ilha de Skye, Escócia [Jagielska et al, 2022]. A região em que este réptil voava era úmida e banhada por águas quentes, onde ele provavelmente se alimentava de peixes e lulas usando suas presas afiadas e dentes bem definidos [Geggel, 2022].
(Crédito da imagem: El fosilmaníaco).
No mar, outros répteis do Jurássico, os ictiossauros,
passaram por um gargalo morfológico após a extinção do final do Triássico, resultando na restrição da exploração de nichos ecológicos e alimentares. Estes répteis marinhos passaram a apresentar plano corporal semelhante ao dos golfinhos, adaptado à predação de perseguição pelágica.
Apresentavam redução dos membros posteriores, órbitas grandes associadas ao mergulho, cauda e nadadeira dorsal semilunares associadas à natação rápida e sustentada [Jamison-Todd et al, 2022].
O ictiossauros se tornaram predadores dominantes e diversos. No Jurássico, dominaram as posições superiores da cadeia alimentar dos ecossistemas marinhos. Muitas espécies de ictiossauros competiam pela mesma variedade de alimentos ou dividiam recursos do seus regimes alimentares, cada uma se especializando em diferentes tipos de presas. A partição destes recursos, além de modos de caça e estilos de natação e sub-habitats oceânicos, permitiu a diversificação dos ictissauros [Jamison-Todd et al, 2022].

Reconstrução do Ophthalmosaurus icenicus.
Os primeiros ictiossauros eram inadequados para explorar águas profundas. Espécies posteriores, como o Ophthalmosaurus icenicus do Jurássico médio e superior, desenvolveram gordura corporal e capacidade de manter seus corpos aquecidos. Embora o vulcanismo marinho tornasse os oceanos mais rasos e mais quentes do que atualmente, em profundidades maiores a água ainda devia ser muito fria [Robinson, 2024].
(Crédito da imagem: Fishboy86164577).
A extinção em massa do final do Triássico influenciou a evolução de outros répteis marinhos, os plesiossauros,
eliminando a maior parte da competição, inclusive dos gigantes ictiossauros. Nos primeiros 20 milhões de anos do Jurássico, com base no registro fóssil europeu, o tamanho máximo do corpo dos plesiossauros evoluiu gradualmente, embora no Jurássico superior houvesse abundância de exemplares pequenos (pesando <100 kg) [Sander, 2023].
Alguns plesiossauros (em que o longo pescoço se tornou mais curto, os dentes mais robustos e o crânio maior) assumiram o estilo macropredatório, alimentando-se de outros répteis marinhos. Os demais plesiossauros (com cabeça pequena, focinho curto e o característico pescoço longo) mantiveram o hábito de se alimentar de peixes. Todos tinham alta capacidade de dispersão e colonizaram toda a Terra devido ao estilo de navegação de baixa energia combinado com endotermia (sangue quente) [Sander, 2023].

Reconstrução do Plesiosaurus dolichodeirus.
O Plesiosaurus dolichodeirus foi encontrado em 1823 pela paleontóloga inglesa Mary Anning (1799-1847) na costa sul da Grã-Bretanha [Bonne, 2024].
(Crédito da imagem: Dmitry Bogdanov).
Provavelmente, usando a recente diversidade de insetos (ver a seguir) como alimento, no Jurássico surgiram os primeiros lagartos [Paselk, 2012]. Destaca-se o Bellairsia gracilis
que vagava no Jurássico médio pelas águas doces da região das atuais Hébridas Interiores, na Escócia, há mais de 166 milhões de anos. Ele apresenta uma mistura de características encontradas em répteis escamatas
antigos e modernos e ajudou a esclarecer algumas relações primitivas entre os lagartos [Ashworth, 2022].
Por outro lado, vivendo em terra firme, os crocodilianos eram pequenos e muito ativos, alguns carnívoros enquanto outros eram herbívoros e onívoros. Com o tempo cresceram e os carnívoros passaram a ser a forma dominante, como os goniofolidídeos,
que surgiram no Jurássico e eram semi-aquáticos. Este estilo de vida foi impulsionado por uma válvula que permitia selar a garganta, enquanto submerso na água, e abrir a boca para atacar suas presas. Isto provavelmente os ajudou a se especializarem como predadores de emboscada [Ashworth, 2021].
Há mais de 150 milhões de anos, o Amphicotylus milesi,
um antigo parente de crocodilos e jacarés modernos, também assumiu estilo de vida semi-aquático. Foi encontrado na Formação Morrison, em Wyoming, EUA, em 1899 [Ashworth, 2021].
4.2.2.6. Aves
Datados desde o Jurássico Médio (até o Cretáceo Inferior), no nordeste da China são encontrados diversos fósseis de dinossauros terópodes com penas. Na Sibéria foi encontrado fóssil de dinossauro com pequenas escamas e estruturas semelhantes a penas sugerindo que escamas e penas poderiam ter coexistido nos primeiros dinossauros. Apêndices epidérmicos semelhantes a cerdas ocorrem também em pterossauros sugerindo que as penas podem ter evoluído independentemente dos terópodes [Godefroit et al, 2014].
Em 1861, foi encontrada na Alemanha a Archaeopteryx,
uma ave primitiva de cauda longa. Era considerada a única ave do Jurássico até surgir na China, em 2023, a Baminornis zhengheis,
também uma ave primitiva, mas de cauda curta. Ambas viviam no Jurássico superior há ~150 milhões de anos. A Baminornis, com ~15 cm de comprimento, era anatomicamente mais avançada e voava melhor do que a Archaeopteryx devido à cauda mais curta. Esta é uma importante inovação aerodinâmica que ajudava a deslocar o centro de massa em direção às asas [Dunham, 2025].

Reconstruções da Archaeopteryx (acima) e da Baminornis zhenghensis (abaixo).
O habitat da Archaeopteryx ficava às margens das lagoas calcárias da atual região de Altmühltal, na Baviera, Alemanha. O clima era quente e úmido e a região era pontilhada por extensas lagoas cercadas por recifes de corais e ilhas. Lama calcária finamente granulada se acumulou entre as estruturas dos recifes, formando os atuais Calcários de Solnhofen, ricos em fósseis como o Archaeopteryx [Senckenberg, 2024].
A Baminornis zhenghensis provavelmente vivia em ambiente pantanoso. A Fauna Zhenghe, onde foi encontrada, é uma biota terrestre com grande número de fósseis de tartarugas, cágados, peixes e plantas. É um local onde fósseis de aves do final do Jurássico estão preservados. Eram tempos de erupções vulcânicas frequentes, mas durante intervalos calmos, lagos e bacias se formaram nas áreas montanhosas do atual Condado de Zhenghe, na China, criando um ambiente ecológico favorável a plantas e animais [Yali, 2025].
A Baminornis zhenghensis provavelmente vivia em ambiente pantanoso. A Fauna Zhenghe, onde foi encontrada, é uma biota terrestre com grande número de fósseis de tartarugas, cágados, peixes e plantas. É um local onde fósseis de aves do final do Jurássico estão preservados. Eram tempos de erupções vulcânicas frequentes, mas durante intervalos calmos, lagos e bacias se formaram nas áreas montanhosas do atual Condado de Zhenghe, na China, criando um ambiente ecológico favorável a plantas e animais [Yali, 2025].
4.2.2.7. Mamíferos
As duas subclasses principais dos mamíferos são constituídas pelos [NHC, 2001]:
- prototérios e
- térios. Estes são subdivididos em:
No Jurássico inferior, a divergência entre os metatérios e eutérios iniciou há ~180 milhões de anos [Kelly e Barr, 2012], quando viviam no alto da copa das árvores em segurança, protegidos dos predadores. A separação foi efetivada há 160 milhões de anos, quando os eutérios placentários,
representados pelo Juramaia sinensis,
seguiram caminhos diferentes dos metatérios marsupiais
[Roach, 2011]. O Juramaia sinensis, portanto, seria o mais antigo ancestral dos placentários, grupo que constitui a grande maioria dos mamíferos na Terra atualmente [Straus, 2019].

Reconstrução do Juramaia sinensis.
No topo das árvores, este mamífero percorria as florestas temperadas do Jurássico, alimentando-se de insetos à noite [Roach, 2011].
(Crédito da imagem: Nobu Tamura)
Apesar de as taxas de evolução morfológica dos mamíferos terem diminuído e permanecerem significativamente baixas até o final do Mesozoico, foram significativamente elevadas antes do Jurássico superior, com um pico ocorrendo durante o Jurássico inferior e médio. Durante o Jurássico médio e superior, os mamíferos experimentaram grande radiação adaptativa
e a explosão intensa de inovação morfológica foi essencial para a origem dos mamíferos do grupo coroa.
Resultou em rápido aparecimento de linhagens anatomicamente distintas em nível de ordem ou família, como triconodontes,
multituberculados
e docodontes
[Close et al, 2015].
Foram gerados altos níveis de diversidades ecológica e morfológica e houve aquisição de características mamíferas importantes em dentição, ouvido médio e cintura escapular. Há duas hipóteses para a motivação de tal evolução: (i) a fragmentação do Pangeia e (ii) a aquisição de uma "massa crítica" de inovações anatômicas ou fisiológicas que vinham se acumulando constantemente desde a origem da linhagem mamífera e antes, nos cinodontes
não mamíferos [Close et al, 2015].
4.2.2.8. Insetos
Os insetos também se diversificaram no Jurássico, surgindo muitas formas modernas como besouros, vespas [Paselk, 2012] e pulgas. Estas originalmente parasitavam os dinossauros emplumados e posteriormente se especializaram em mamíferos e aves. No Jurássico superior, os cupins inauguraram as primeiras sociedades de insetos, um sistema apoiado pela construção coletiva de um ninho [Engel, 2015].

Reconstrução da pulga Pseudopulex jurassicus,
imaginada parasitando um dinossauro emplumado [Poinar, 2012]
(Crédito da imagem: Wang Cheng).
4.2.2.9. Anfíbios
Girinos e adultos do anfíbio Notobatrachus degiustoi
[Buehler, 2024] foram encontrados na Patagônia da Argentina. Seus girinos (larvas de anfíbios anuros)
são os mais antigos conhecidos no registro fóssil. Este sapo, em seu ciclo de vida bifásico (com fases de girino e adulta), apresenta girinos que se alimentam por filtragem habitando ambientes aquáticos efêmeros. Remontando ao Jurássico médio-superior (~161 milhões de anos atrás), o Notobatrachus degiustoi revela a história evolutiva inicial dos anuros ancestrais e seu tamanho demonstra que o gigantismo já ocorria entre eles. Seus estágios de metamorfoses com profundas mudanças morfológicas e ecológicas até atingir idade reprodutiva adulta são os mais extremos entre os tetrápodes [Chuliver et al, 2024].

Reconstrução do Notobatrachus degiustoi.
Foi encontrado em 2020 na Formação La Matilde, na Província de Santa Cruz, Argentina, cujos sedimentos foram depositados em planície de inundação. Era um ambiente de baixa energia de lagoas, lagos ou pântanos, sob clima sazonal que variava entre seco e úmido. Naqueles sedimentos também foram encontrados fósseis de plantas, crustáceos braquiópodes, bivalves e insetos [Chuliver et al, 2024].
(Crédito da imagem: Nobu Tamura).
4.2.2.10. Peixes
No caso dos peixes, os teleósteos,
evoluídos durante o Jurássico há mais de 150 milhões de anos, foram os que mais dominaram os mares, tanto em habitats marinhos quanto de água doce, dos trópicos às regiões polares. Pertencem a classe Actinopterygii
e incluem a maioria dos peixes atuais, como salmão, atum, bagre, bacalhau e muitos outros [basicbiology, 2015].
No Jurássico inferior, há ~195 milhões de anos, os tubarões Hexanchiformes
desenvolveram mandíbulas flexíveis e salientes, permitindo que se alimentassem de presas maiores do que eles. Também passaram a ter a capacidade de nadar mais rapidamente. No restante do Jurássico, foram seguidos pela maioria dos grupos modernos de tubarões [Davis, 2025].
As raias (ou arraias)
tiveram grande diversidade no Jurássico superior. Um exemplo é a Aellopobatis bavarica,
encontrada no Arquipélago Solnhofen, na Baviera, Alemanha [Türtscher et al, 2024].

Foi encontrado em calcário marinho de granulação fina do Jurássico superior, chamado "plattenkalk" [Türtscher et al, 2024; Türtscher et al, 2025]. Estes calcários se formam em ambientes carbonáticos protegidos da atividade de ondas ou correntes, onde a água frequentemente apresenta salinidades anormais e baixas concentrações de oxigênio [Swinburne e Hemleben, 1994].
(Crédito da imagem: Türtscher et al).
4.2.2.11. Plantas
O Jurássico também é frequentemente chamado de "Era das Cicadáceas" porque elas constituíram 20% da flora da Terra quando (Triássico e Jurássico) apresentavam sua maior diversidade. As cicadáceas coexistiram com os dinossauros e provavelmente foram consumidas por eles. Ao contrário dos dinossauros, porém, não se extinguiram, embora não sejam tão abundantes ou diversas como no Mesozoico [UCMP, 2025]. No Jurássico inferior, na região da atual Europa, as cicadáceas foram relevantes sendo a Nilssonia acuminata
a espécie mais difundida [Wachtler, 2024a].

Fóssil de Nilssonia encontrado no Membro Stanstorp do Arenito Höör, no condado de Skåne, no sul da Suécia, em exposição no Museu de História Natural de Viena, Áustria.
O Arenito Höör apresenta conjuntos palinológicos
que revelam ecossistemas associados a habitats com perturbações climáticas [Vadja et al, 2016]. Este arenito, do Jurássico Inferior da Suécia, é subdividido em dois membros. O Membro Stanstorp representa sedimentos fluviais com uma camada conglomerática intercalada representando um forte evento de inundação. O Membro Vittseröd sobreposto foi depositado em uma plataforma marinha dominada por tempestades e marés [Pieńkowski, 2002].
(Crédito da imagem: Gyik Toma).
As cicadáceas eram um componente marcante da vegetação em número de espécies, em tamanho e em quantidade de indivíduos. Árvores e arbustos de cicadáceas, juntamente com coníferas e ginkgoales,
dominaram as florestas do Mesozoico. As cicadáceas se estenderam pela Terra desde o Alasca e a Sibéria até a Antártida. O clima geralmente mais quente e com maior umidade em comparação com o atual permitiu que elas fossem encontradas em todos os continentes [UCMP, 2025], ou seja, a abundância de cicadáceas é consistente com o clima subtropical que prevalecia [Wachtler, 2024a].
Quanto às coníferas, durante o início do Jurássico, abrangendo regiões da Europa ao Quirguistão, China e Mongólia, três gêneros destas plantas dominaram grandes áreas do hemisfério norte: Podozamites distans,
Swedenborgia liaso-keuperianus
e Hirmeriella muensteri.
O primeiro, o mais difundido, provavelmente perdia suas folhas sazonalmente, conforme indicam extensos tapetes de folhas fósseis. O terceiro se destaca como uma conífera característica e facilmente distinguível no Jurássico da Baviera [Wachtler, 2024b]. As coníferas dominavam florestas produtoras de carvão [Paselk, 2012], entretanto, atualmente, muitas estão confinadas a zonas relíquias [Wachtler, 2024b].
4.2.2.12. A tectônica nos oceanos Pacífico e Atlântico
No Jurássico superior, os padrões paleogeográficos foram influenciados pela ruptura do Pangeia e, também, pelo crescimento da placa tectônica do Pacífico. Tais padrões impactaram o clima global que se caracterizava por um efeito estufa quente e uniforme [Wang et al, 2024].
Acredita-se que o Oceano Pantalassa, que circundava o Pangeia, hospedou pelo menos três placas tectônicas, todas compartilhando cristas em expansão com a crescente Placa do Pacífico. Eram a conceitual Placa Phoenix ao sul, a Placa Farallon a nordeste e a Placa Izanagi a noroeste [Boschman et al, 2021]. A reorganização da expansão da junção tripla Pacífico-Izanagi-Farallon no Jurássico superior produziu a Elevação Shatsky
localizada na bacia noroeste do Pacífico sobre o fundo marinho [Nakanishi et al, 2015]. O estudo da formação de planaltos oceânicos, como a Elevação Shatsky, ajuda a entender eventos anóxicos oceânicos e extinções em massa que ocorrem associados a eles [Tominaga Hara, 2021].

Localização atual (nos quadros com bordas brancas) da Elevação Shatsky e da cadeia havaiana dos Montes Submarinos Imperador.
(Imagem baseada em: Seattle Skier; mlp.ldeo).
O Oceano Pacífico ocupava quase um hemisfério inteiro e, à medida que se estreitava, o Atlântico se alargava, assim como ocorre atualmente. Durante o Jurássico superior (e o Cretáceo), as conexões entre estes dois oceanos, por meio de passagens entre pequenas placas continentais na América Central, ampliavam-se gradualmente, viabilizando um sistema de correntes que fluía principalmente para oeste em baixas latitudes ao circundar a Terra [Winterer, 1991].
A expansão do fundo do mar e a criação do Oceano Atlântico Norte resultou a partir da formação de uma série de bacias sedimentares delimitadas por falhas. Algumas foram formadas devido à fragmentação do Pangeia, iniciada na fronteira Triássico-Jurássico, e à extensão entre os escudos Laurentiano, Báltico e Avaloniano [Morton et al, 2024].
Um ciclo de rifting, associado à evolução das bacias intraplacas ibéricas, foi retomado no início do Jurássico superior com três pulsos discretos que controlaram o desenvolvimento daquelas bacias que faziam parte da plataforma noroeste que circundava o Tétis [Salas et al, 2010]
Tais bacias de rifte formaram-se entre a Europa e a América do Norte/Groenlândia, com ligações variáveis aos oceanos Tétis e Ártico. Os sistemas de rifte Ártico e proto-Atlântico permaneceram separados pelo menos até o Jurássico superior. Os sedimentos jurássicos nessas bacias de rifte posteriormente forneceram rochas-fonte para produção de reservatórios importantes com recursos significativos de hidrocarbonetos [Morton et al, 2024].
4.2.2.13. Hidrocarbonetos
No Jurássico, em certos momentos houve grandes surtos de atividade vulcânica que desencadearam concentrações crescentes de CO₂ atmosférico, elevação da temperatura da superfície do mar e redução dos níveis de oxigênio em grandes áreas dos oceanos. Assim, os níveis de nutrientes aumentaram no ambiente marinho ocasionando proliferação de algas e bactérias. A morte destes organismos levou ao acúmulo de matéria orgânica preservada em sedimentos depositados no fundo do mar [Butterfield, 2017].
A abundância de matéria orgânica em rochas sedimentares, como no caso do Jurássico, é um dos requisitos para potencializar a geração de hidrocarbonetos
e a formação de depósitos de óleo e gás [Deng et al, 2025]. Estima-se que o Jurássico superior representa 25% das reservas mundiais de petróleo, abrangendo as bacias da Arábia-Irã, da Sibéria Ocidental, do Golfo do México e do Mar do Norte [Peters et al, 2008].
Eis alguns exemplos de depósitos de petróleo e gás do Jurássico:
- A Bacia Intraplataforma Arábica (Jurássico médio e superior) constitui cenário de uma das maiores reservas convencionais de petróleo da Terra. Nesta bacia está localizado o Campo Ghawar, considerado o maior campo de petróleo do mundo [Wilson, 2020].
- Folhelhos da Argila Kimmeridge e equivalentes (Jurássico superior a Cretáceo inferior), principais rochas-fonte do petróleo em Graben do Mar do Norte, foram depositadas do Jurássico superior ao Cretáceo inferior durante intensa extensão e rifteamento. As rochas geradoras têm sido continuamente soterradas desde a deposição. A formação de traps estruturais também ocorre continuamente desde o Jurássico superior. Como resultado, petróleo e gás estão presentes em uma ampla variedade de cenários [Gautier, 2005].
- As formações Sargelu e Naokelekan são as principais rochas-fonte de petróleo no Curdistão iraquiano. A rocha geradora de Sargelu (Jurássico médio) é caracterizada por alto teor de matéria orgânica e enxofre [Hakimi et al, 2018]. A Formação Naokelekan (Jurássico médio e superior) é uma sequência heterogênea composta principalmente por folhelhos betuminosos com calcário betuminoso e calcários dolomíticos [Abdula, 2016].

Localizações atuais da Bacia Intraplataforma Arábica (na área com bordas vermelhas) e de afloramentos do Jurássico na Península Arábica.
4.2.3. Cretáceo
| De 145 milhões a 66 milhões de anos atrás | |
| Luminosidade do Sol | de 98,7 a 99,3% da atual |
| Duração do ciclo dia-noite | de 23,1 a 23,6 horas |
| Temperatura | de 27,2 a 25,2°C com mínima de 24,4 e máxima de 35,8°C |
| Concentração de O₂ na atmosfera | de 15,2 a 22% |
| Quantidade de CO₂ na atmosfera | de 540 a 614 ppm com máxima de 1123 ppm |
4.2.3.1. Deriva continental e vulcanismo
A tectônica, a geografia e o clima no Cretáceo eram muito diferentes dos atuais. No início deste período, quando o Pangeia se fragmentava, apenas algumas pequenas bacias oceânicas separavam Laurásia, Gondwana Ocidental e Gondwana Oriental. Nenhuma grande cadeia de montanhas colisional entre continentes se formou no Cretáceo. Quase todas as montanhas foram formadas ao longo das margens de subducção dos oceanos Pantalassa e Tétis. O interior dos continentes era em grande parte plano, com exceção da Ásia Central, onde cadeias de montanhas do Paleozoico superior ainda se destacavam na paisagem. À medida que os continentes se separavam, surgiam bacias oceânicas jovens, onde a litosfera termicamente elevada impulsionava o nível do mar, mais alto que o atual, que inundava os continentes com efeitos globais importantes no clima [Scotese et al, 2024].
A ruptura de Pangeia foi acompanhada por extensa atividade magmática episódica [Peace et al, 2020]. A sequência de rifteamento passivo/ativo observada desde o Carbonífero continuou no Cretáceo [de Lamotte et al, 2015]. O rifteamento começou no sul no final do Jurássico e atingiu a zona equatorial no final do Cretáceo [Torsvik et al, 2009].
Os répteis crocodilianos Sarcosuchus imperator
e Sarcosuchus hartti,
encontrados, respectivamente, no sítio de Gadoufaouna, no Níger (ao norte da Bacia do Gabão), África, e na Bacia do Recôncavo, na Bahia, Brasil, América do Sul, são ambos do Cretáceo inferior. Eles testemunharam a fragmentação do Gondwana com a abertura do Oceano Atlântico Sul e, antes disto, seu ancestral comum, do gênero Sarcosuchus, testemunhou a união da América do Sul com a África [Taquet, 2021].

Reconstrução do Sarcosuchus imperator que muito bem poderia ser do Sarcosuchus hartti, devido às semelhanças entre eles.
A Bacia do Recôncavo, na Bahia, tem sua parte oriental na costa ocidental da África, onde a Bacia do Gabão apresenta uma sucessão de depósitos semelhantes e igualmente de ambiente de água doce. A formação do graben do Recôncavo-Gabão, no início do Cretáceo, é considerada um estágio inicial na separação entre a África e a América do Sul. A presença dos crocodilianos não marinhos do gênero Sarcosuchus, tanto no Brasil quanto no Níger, fornece evidências adicionais da continuidade faunística entre as duas regiões antes da abertura do Oceano Atlântico Sul [Taquet, 2021].
(Crédito da imagem: Rhunevild)
O Atlântico Sul começou a se abrir a partir da erupção de um vulcanismo do tipo hotspot
na Dorsal Mesoatlântica.
Este vulcanismo formou duas grandes províncias ígneas do Atlântico Sul: a Elevação do Rio Grande,
na placa Sul-Americana, e a Dorsal Walvis,
na placa Africana. Embora não constituam um único lineamento devido à reorganização das elevações e à formação de microplacas, são consideradas semelhantes à clássica cadeia dos Montes Submarinos Imperador (oriunda também de hotspot na região do Havaí) [Sager et al, 2021].

Localizações atuais da Elevação do Rio Grande e da Dorsal Walvis (nos quadros com bordas brancas), da Dorsal Mesoatlântica, das ilhas Tristão da Cunha e Gonçalo Álvares (ou Gough) e da Província Ígnea Paraná-Etendeka (em vermelho).
A tectônica distensiva que causou a ruptura do Gondwana gerou o intenso vulcanismo da Província Ígnea Paraná-Etendeka
[Benites et al, 2020] associado ao hotspot sob as ilhas Tristão da Cunha e Gonçalo Álvares. Os volumosos derrames basálticos continentais desta província se distinguem por rápida erupção, principalmente entre 133 milhões e 130 milhões de anos. Na América do Sul (Paraná), as lavas cobrem uma área de ∼1,2-1,6 × 10⁶ km², enquanto no lado africano (Etendeka), cobrem uma área de ∼0,8 × 10⁵ km². Lavas associadas estão espalhadas ao longo de mais de 1.500 km da costa da África Ocidental [Torsvik et al, 2009].
A crosta oceânica inicial do Atlântico Sul foi gerada durante o Supercrono Normal do Cretáceo
[Torsvik et al, 2009]. As taxas de expansão do fundo do mar aumentaram de forma constante durante a maior parte do Cretáceo e diminuíram acentuadamente há ~80 milhões de anos [Granot e Dyment, 2015].
Outras fragmentações ocorreram, por exemplo, com a Índia, que se transformou em um continente insular [USGS, 2025]. As separações entre Índia e Madagascar e entre Austrália e Antártida ocorreram no Cretáceo superior [de Lamotte et al, 2015]. A partir deste conjunto de fragmentações, os oceanos atingiram seus níveis mais elevados, resultando em vastos mares continentais rasos. A partir de restos esqueléticos de organismos marinhos, enormes depósitos de giz surgiram dando nome ao período Cretáceo [Paselk, 2012].
No Cretáceo Superior, a maioria das placas tectônicas se moveram para suas posições atuais enquanto o nível do mar continuou a subir impulsionado em parte pelos processos tectônicos. Isto levou a extensas inundações de regiões baixas e a uma diminuição substancial na área terrestre que ficou ainda mais fragmentada [Vavrek, 2016]. Com a elevação do nível do mar e a separação dos continentes, as correntes oceânicas rasas levaram a água quente para mais longe em direção aos polos, o que propiciou um clima global com polos sem gelo [USGS, 2025].
4.2.3.2. Clima quente
A tendência de aquecimento do clima que já vinha acontecendo no Jurássico perdurou no Cretáceo [Scotese et al, 2024] com condições quentes presentes e até calor extremo alcançado no Máximo Térmico do Cretáceo
por volta de ~91 milhões de anos [Poulsen, 2025]. Após, um resfriamento global de longo prazo teve início há ~83 milhões de anos e culminou em mínimos de temperatura no final do Cretáceo [Huber et al, 2018].
Durante todo o período ocorreu um aumento relativamente maior das temperatura em direção aos polos [Vavrek, 2016], sem permitir grandes calotas de gelo permanentes, embora pequenas calotas (~4 × 10⁶ km²) possam ter existido no Polo Sul no início e no final do Cretáceo [Scotese et al, 2024].

Configuração dos continentes da Terra há 105 milhões de anos com sobreposição dos contornos dos países atuais em suas respectivas localizações.
Reconstrução paleogeográfica com altimetria desde azul escuro (oceanos profundos) até azul claro (litosfera continental inundada e ilhas oceânicas) e desde verde (terras baixas) até marrom (montanhas) [Scotese et al, 2024].
É provável que oceanos quentes que cercavam e penetravam o interior da Antártida (devido ao nível do mar relativo mais alto) tenham impedido o crescimento de camadas de gelo na região [Huber et al, 2018].
(Crédito da imagem: Scotese et al).
Há indícios de tendências de temperaturas semelhantes nas águas do fundo e da superfície tanto no sul do Atlântico Sul quanto nas bacias do Oceano Índico. Plantas terrestres e dinossauros de latitudes polares de ambos os hemisférios indicam temperaturas médias anuais e mínimas de inverno muito mais altas do que as atuais. A presença de champsossauros
em regiões com latitude 72°N (entre ~93 milhões e ~87 milhões de anos) é consistente com temperaturas médias anuais >14°C naquelas regiões [Huber et al, 2018].
O clima de estufa quente do Cretáceo é explicado pelos elevados níveis de CO₂ atmosférico devido à liberação de gases vulcânicos. Os efeitos de contrapeso do aumento do intemperismo químico de rochas de silicato e do aumento do soterramento de carbono orgânico podem também ter tido influência na elevação das temperaturas [Huber et al, 2018].
O acúmulo de CO₂ atmosférico, a paleogeografia e os oceanos quentes do Cretáceo criaram condições próximas ao limiar para a deposição generalizada de sedimentos ricos em matéria orgânica sob condições anóxicas. Isto é demonstrado pela ocorrência repetida de eventos anóxicos oceânicos. Estes eventos do Cretáceo com expressão global ocorreram há ~120 milhões e ~94 milhões de anos. Regionalmente, ocorreram há ~111 milhões, ~100 milhões e possivelmente ~86 milhões de anos. Há propostas de que estes eventos sejam explicados pela redução da circulação oceânica (devido a bacias oceânicas estagnadas), pela elevada produtividade do plâncton e pela expansão da zona mínima de oxigênio [Huber et al, 2018].
A alteração na geografia e a tendência de aquecimento favoreceu a biodiversidade. A fragmentação dos continentes promoveu também o desenvolvimento de regiões com conjuntos faunísticos distintos e, inclusive, com endemismos.
Nos últimos 5 milhões de anos do Cretáceo, o nível do mar começou a cair, reconectando regiões anteriormente separadas, levando a uma homogeneização prevista dos ecossistemas e à diminuição geral da biodiversidade [Vavrek, 2016].
4.2.3.3. A Revolução Terrestre do Cretáceo e as angiospermas
Há ~110 milhões da anos, a biodiversidade marinha foi superada pela terrestre que explodiu 10 milhões de anos depois na chamada Revolução Terrestre do Cretáceo (especialmente entre 145 milhões e 66 milhões de anos atrás). Esta explosão coincidiu com inovações na biologia das angiospermas
[Benton e Sauquet, 2021], cuja biodiversificação acelerada no Cretáceo médio foi notada inclusive pelo naturalista, geólogo e biólogo britânico Charles Robert Darwin (1809-1882), que a registrou em carta de 1879 [Birks, 2020]. Esta biodiversidade incluiu [Benton e Sauquet, 2021]:
- o aparecimento das flores, que tornaram a reprodução mais eficiente;
- a coevolução com animais, especialmente polinizadores e herbívoros;
- a maior capacidade de fotossíntese; e
- a adaptabilidade e capacidade de modificar habitats.
Nestes tempos, dois Eventos Anóxicos Oceânicos, no Cretáceo Inferior e Superior, impactaram fortemente a vida marinha e mais fracamente os ecossistemas terrestres. Neste contexto aconteceu a evolução das angiospermas, que passaram a superar outras plantas em número de espécies [Benton e Sauquet, 2021], principalmente as gimnospermas e, dentre estas, particularmente as coníferas [Birks, 2020].
Fizeram parte da Revolução Terrestre do Cretáceo [Benton e Sauquet, 2021]:
- Fungos – Novos fungos se diversificaram associados à explosão das angiospermas e, por sua vez, forneceram água e nutrientes minerais às plantas terrestres.
- Insetos – Calcula-se que 80% das espécies atuais de insetos pertencem a famílias que surgiram no Cretáceo (com pico em 125 milhões de anos atrás), o que é geralmente explicado pelo aumento de suas relações coevolutivas com as angiospermas. São incluídos nestas relações insetos como besouros, gafanhotos, grilos, percevejos, cigarras, borboletas, mariposas, baratas, moscas, vespas e abelhas. Exemplos:
- Baratas – Podem ter tido novas oportunidades proporcionadas pelo aumento geral nos níveis de nitrogênio nos solos decorrente da produtividade da angiospermas.
- Moscas – Alimentavam-se de angiospermas. Restos de grãos de pólen de angiospermas são encontrados em intestinos de moscas fósseis.
- Vespas e abelhas – São relacionadas à angiospermas através da polinização.
- Aracnídeos – Exemplos:
- Aranhas – Como predadoras de insetos, diversificaram-se entre 125 milhões e 100 milhões da anos atrás.
- Ácaros – Surgiram no Cretáceo superior e viviam sob as folhas das angiospermas. Alimentavam-se de fungos e presas e davam proteção à planta.
- Tetrápodes – Vários grupos de vertebrados modernos se diversificaram no Cretáceo. Há indícios de algumas conexões plausíveis com a explosão das angiospermas. Exemplos:
- Anfíbios – Viviam em florestas tropicais úmidas dominadas por angiospermas. Alimentavam-se de insetos, aranhas, caracóis, lesmas e minhocas.
- Aves – Há evidências também de que aves do Cretáceo se alimentavam de angiospermas.

Pticopterídeo macho
preservado em âmbar birmanês
do Cretáceo (~100-110 milhões de anos atrás) parasitado por larvas de ácaro, aderidas predominantemente ao abdômen do inseto.
O âmbar birmanês é fonte significativa de fósseis de plantas e animais que documentam o bioma do Cretáceo médio do sudeste da Ásia, mostrando evidências de ecossistema tropical quente. São encontradas espécies terrestres e marinhas, sugerindo que as árvores (de onde se originou o âmbar) estavam próximas a habitats estuarinos e de água doce. Nestes tempos, a região se separou do Gondwana e, após, colidiu com a Ásia [Wood e Wunderlich, 2023].
(Crédito da imagem: Brochard).
A contribuição das angiospermas para a Revolução Terrestre do Cretáceo provavelmente se deve às adaptações desenvolvidas por elas. Tais adaptações permitiram que superassem as restrições que limitavam a diversidade de espécies e dos nichos ecológicos de outras plantas terrestres. Foram adaptações graduais que ocorreram com temperaturas globais elevadas primeiramente flutuantes e posteriormente em declínio em continentes fragmentados. Assim, clima e paleogeografia impulsionaram muitos aspectos da evolução das angiospermas [Benton e Sauquet, 2021].
Acredita-se que o domínio das angiospermas tenha alterado fortemente o clima e os ciclos hídricos. Tais alterações estão associadas às elevadas capacidades das angiospermas de aumentar o intemperismo do substrato rochoso, absorvendo assim o CO₂, e de transpirar água extraída do solo, passando-a à atmosfera. Estes efeitos são mais evidentes nas florestas tropicais úmidas [Benton e Sauquet, 2021].
4.2.3.4. Reestruturação das comunidades bentônicas no Cretáceo
Esta reestruturação (ou revolução) que já vinha acontecendo no Jurássico, continuou por todo o Cretáceo como um processo evolutivo percebido como uma série de eventos, sendo muitos deles não relacionados entre si. Estes eventos ocorreram como resposta coevolutiva das presas aos predadores durófagos (como já foi visto no Jurássico). Entre as consequências, continuou havendo um aumento substancial da robustez das conchas dos bivalves e a transformação das comunidades bentônicas marinhas assumindo a infaunalização [Lucas e Hunt, 2023].
No Cretáceo, a diversidade de equinoides com infaunalização aumentou e é vista como um dos eventos evolutivos mais importantes na história deste grupo. No caso dos moluscos, tendências de infaunalização são fortemente correlacionadas com a intensificação da predação com perfuração de conchas [Petsios et al, 2021].
Portanto, as tendências do Jurássico tiveram continuação no Cretáceo, embora no final deste período um padrão de acoplamento bentônico-pelágico de estilo moderno já tenha sido estabelecido acompanhado por um aumento na icnodiversidade global.
O rápido desenvolvimento de cocolitóforos
forneceu um novo substrato (giz) [Buatois et al, 2016]. Suas ações formaram esqueletos de calcita e seus subsequentes afundamentos no oceano modificaram a alcalinidade da água de pouca profundidade e afetaram diretamente a troca ar/mar de CO₂. [Rost e Riebesell, 2021]. Além disto, o fluxo crescente de matéria orgânica para o fundo do mar permitiu o estabelecimento de estruturas de estratificação complexas, desconhecidas anteriormente. O grau de macrobioerosão
indicou uma participação crescente de alguns agentes típicos de comunidades modernas [Buatois et al, 2016].

Afloramentos de giz do Membro Smoky Hill da Formação Niobrara, nas proximidades de Castle Rock, no oeste de Kansas, EUA.
O Membro Smoky Hill foi depositado na parte oriental de uma ampla via marítima, no Cretáceo superior, que se estendia do Golfo do México ao Canadá Ártico. O principal componente da maioria dos seus estratos é carbonato de cálcio, principalmente calcita com baixo teor de magnésio. Consiste em grande parte de restos esqueléticos de cocolitóforos (KGS, 2015).
(Crédito da imagem: James St. John).
4.2.3.5. Mamíferos
O mais antigo mamífero monotremado
conhecido, o Teinolophos trusleri,
encontrado no Cretáceo inferior, foi recentemente alinhado com os ornitorrincos.
Em estudos anteriores, porém, ele foi considerado um monotremado primitivo, anterior à separação entre ornitorrincos e equidnas.
Estes dois grupos de mamíferos pequenos posteriormente aumentaram o tamanho corporal, alcançando ~700 g a ~18 kg de peso [Phillips et al, 2009]. Também do Cretáceo inferior é o Steropodon galmani,
um monotremado semelhante a ornitorrinco que viveu ao lado de dinossauros, crocodilos, aves primitivas e outros mamíferos primitivos [Musser, 2024]

Reconstrução do Steropodon galmani, um monotremado do Cretáceo inferior da Austrália.
Viveu nas margens florestadas do Mar de Eromanga, próximas ao Círculo Polar Antártico, há 110 milhões de anos. Havia coníferas araucárias como árvores dominantes e o sub-bosque provavelmente incluía samambaias e cicadáceas [Musser, 2024].
(Crédito da imagem: Nobu Tamura).
Ainda enquanto mamíferos pequenos, o tamanho corporal, a dieta e a locomoção de comunidades destes animais no Cretáceo superior estão relacionados às condições de recursos locais versus diversidade e também à riqueza e à disparidade ecológicas de tais comunidades. Acredita-se que a evolução dentária deles, o surgimento das angiospermas e a competição com outros vertebrados foram cruciais na formação da estrutura ecológica das comunidades de mamíferos de corpo pequeno ao longo do tempo [Chen et al, 2019].
No final do Cretáceo, a evolução da anatomia foliar das angiospermas, com maiores taxas de transpiração de água, teria proporcionado habitats não arbóreos (no chão) em ambientes de sub-bosque mais complexos e com maior diversidade de recursos alimentares (vegetais e invertebrados) para os mamíferos. Este fato pode estar associado à seletividade das extinções do final do Cretáceo, quando mamíferos metatérios (marsupiais) arbóreos tiveram níveis mais elevados de extinção que mamíferos eutérios (placentários) não arbóreos [Janis et al, 2025]
4.2.3.6. Répteis
Os répteis tiveram seu ápice no Cretáceo, principalmente os dinossauros e os pterossauros. [Scotese et al, 2024]. Já no final do Jurássico, os pterossauros com cauda longa foram praticamente substituídos pelos de cauda curta, como o pterodáctilo
e o pteranodonte.
Com asas maiores e mais manobráveis, eram capazes de planar mais longe, mais rápido e mais alto no céu, descendo como águias para pescar em oceanos, lagos e rios [Strauss, 2019]. No caso dos plesiossauros, os maiores podiam atingir 6 toneladas, como os elasmossauros,
que apresentavam pescoço longo desproporcional ao corpo [Sander, 2023].
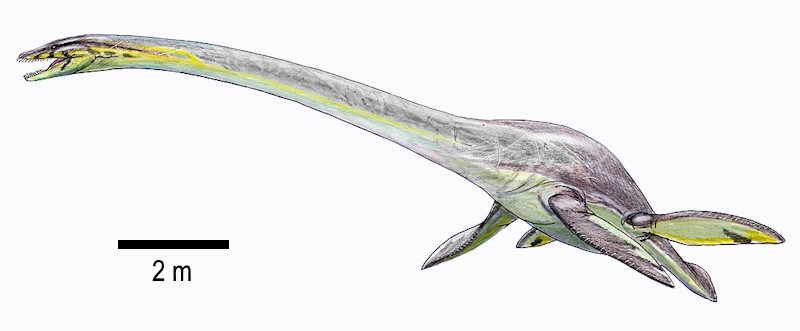
Foi encontrado no Membro Sharon Springs do Folhelho Pierre, Kansas, EUA. Sharon Springs é um folhelho negro rico em matéria orgânica depositado em ambiente anóxico durante transgressão marinha [Carpenter, 1980].
(Crédito da imagem: Dmitry Bogdanov).
Apesar das profundas adaptações ao ambiente aquático e do aparente sucesso mantido ao longo do Cretáceo Inferior, os ictiossauros foram extintos cerca de 30 milhões de anos antes da extinção em massa do final do Cretáceo [Fischer et al, 2016]. Seu declínio está relacionado a um período de instabilidade climática ocorrido há ~95 milhões de anos [Jamison-Todd et al, 2022]. Houve alterações ambientais globais resultando em grande evento de renovação temporalmente escalonado que reorganizou profundamente os ecossistemas marinhos. Uma parte substancial da diversidade dos ictiossauros desapareceu, eliminando Ophthalmosaurus e a maior parte da diversidade ecológica que estava presente no final do Cretáceo Inferior. Foram tempos peculiares, representando o ápice de numerosas perturbações climáticas oceânicas, sem gelo polar, níveis do mar extremamente altos, sedimentação única e forte anoxia, além de temperatura e níveis de CO₂ muito altos [Fischer et al, 2016].
4.2.3.7. Aves
E surgiram as aves mergulhadoras. No Cretáceo superior, as Hesperornithiformes,
primeiras aves conhecidas a se adaptarem a um estilo de vida totalmente aquático, eram mergulhadoras praticamente incapazes de voar. Eram amplamente distribuídas pelo Hemisfério Norte com diversificação relativamente rápida em uma ampla gama de tamanhos corporais e graus de adaptação à água, desde as pequenas Enaliornis
e Pasquiaornis,
com menores graus de especialização em mergulho, até as grandes Hesperornis,
com extremas especializações morfológicas [Bell e Chiappe, 2022].

Reconstrução da Hesperornis.
A ampla distribuição biogeográfica de fósseis da Hesperornis em depósitos do Mar Interior Ocidental
no Cretáceo superior levantou questões sobre se elas suportavam invernos polares ou migravam entre latitudes médias e altas. A Hesperornis tem registro fóssil no Hemisfério Norte, desde médias latitudes até o Ártico canadense. Esta extensa distribuição biogeográfica indica que foram bem-sucedidas em uma grande variedade de ambientes. No Cretáceo superior, as temperaturas da superfície do mar ao longo do Mar Interior Ocidental são estimadas entre 22 e 24°C no Golfo do México e entre 8 e -5°C no Ártico [Wilson e Chin, 2014]
(Crédito da imagem: UDL).
4.2.3.8. Dinossauros
Antes das aves mergulhadoras, os microraptores
ajudaram a estabelecer a evolução das aves a partir dos dinossauros já no Cretáceo inferior. Pesando entre 900 g e 1,3 kg, foram os menores dinossauros e eram emplumados semelhantes a um pássaro. Como característica marcante, possuíam quatro asas, sendo duas nos membros anteriores e duas nos posteriores. Esta característica os tornavam corredores desajeitados, aumentando a suspeita de que podiam no máximo alçar voo, provavelmente saltando de galho em galho nas árvores [Strauss, 2029].

Reconstrução do Microraptor zhaoianus.
Partes do corpo de um lagarto foram encontrados no abdômen de um Microraptor zhaoianus na Formação Jiufotang, na China [O'Connor et al, 2019]. Conforme indicam os sedimentos daquela formação e a matéria orgânica depositada neles, presa e predador lutavam pela sobrevivência em ambiente do Cretáceo inferior onde o clima sofria intensas alterações, passando de relativamente seco para úmido [Liu et al, 2024].
(Crédito da imagem: Entelognathus).
Em meados do Creáceo superior, prosperaram os velociraptores,
que eram dinossauros dromaeossaurídeos.
Eram tempos de mudanças e diversificações significativas no mundo dos dinossauros, com novas espécies surgindo e antigas sendo extintas. Os velociraptores não eram dinossauros grandes, mas eram predadores temíveis equipados com ferramentas e adaptações necessárias para sobreviver em ambiente hostil e competitivo. Suas adaptações, que incorporavam uma mistura única de agilidade, astúcia e puro instinto predatório, transformaram os velociraptores em elementos relevantes no ecossistema dinâmico em que viviam [Douglas, 2024].
São conhecidas duas espécies de velociraptores: mongoliensis e osmolskae,
a primeira foi encontrada na Mongólia e nomeada em 1924 e a segunda foi encontrada na China e nomeada em 2008 [Hendry, 2023].

Reconstrução do Velociraptor mongoliensis.
Apesar da plumagem semelhante à das aves, os velociraptores não voavam pois tinham membros anteriores curtos [natgeo, 2023].
(Crédito da imagem: Fred Wierum).
No Cretáceo superior, com clima global quente, a exuberante vida vegetal sustentava uma fauna diversificada, incluindo insetos, mamíferos e dinossauros [NMHN, 2014]. Neste cenário, reinava o mais temível carnívoro de todos os tempos, o dinossauro Tiranossauro rex,
no oeste da atual América do Norte [AMNH, 2025]. Os tiranossaurídeos,
embora tenham surgido há ~170 milhões de anos, permaneceram pequenos e somente há ~80 milhões de anos começaram a atingir tamanhos gigantescos [Black, 2021].

O primeiro fóssil de Tiranossauro rex foi encontrado em 1902 na Formação Hell Creek, em Montana, EUA [AMNH, 2025]. Nesta região, o clima do Cretáceo superior era temperado quente a subtropical e as florestas, lagoas e pântanos forneciam habitats abundantes para vários tipos de animais, incluindo o T. rex [Hoganson, 2012].
(Crédito da imagem: DBCLS).
Devido ao tamanho do corpo e à força da mordida, os tiranossauros, desde bebês, preencheram muitos nichos em seus ambientes, dominando de forma tirana os ecossistemas do Cretáceo. Eles afastavam outros carnívoros, criando situações diferentes de tudo o que se vê hoje, com ecossistemas dominados por um único grande predador [Black, 2021]. Um tiranossauro adulto podia exercer uma força de mordida capaz de estilhaçar ossos, graças aos músculos expandidos da mandíbula e a um crânio e dentes robustos [Johnson-Ransom et al, 2023].
Por falar em robustez, Titanossauros,
como o Puertasaurus reuili,
o Argentinosaurus huinculensis
e o Patagotitan mayorum,
são de longe os maiores animais do nosso planeta que já viveram em ambiente terrestre [Hendry, 2023]. Estes dinossauros herbívoros quadrúpedes bem-sucedidos no Cretáceo, distribuíram-se amplamente em todos os continentes e em diferentes latitudes, mas principalmente no Gondwana e especialmente na América do Sul [Fiorelli et al, 2022]. Foram os últimos saurópodes antes da extinção em massa do final do Cretáceo [Strauss, 2020].
Seu sucesso pode ter ocorrido por vários fatores fisiológicos, ecológicos e características morfológicas adquiridas ao longo de sua história evolutiva, porém, o comportamento generalista de nidificação, utilizando diferentes paleoambientes e estratégias, foi fundamental. Sítios de nidificação de titanossauros foram encontrados extensivamente ao redor do mundo, mas 75% das espécies de titanossauros foram registradas na Patagônia, Argentina [Fiorelli et al, 2022].

Reconstrução do Patagotitan mayorum.
Este titanossauro foi encontrado nas proximidades de La Flecha, na Patagônia, Argentina, no sítio arqueológico que faze parte do Membro Cerro Castaño da Formação Cerro Barcino [Douglas, 2023]. Esta formação produziu uma notável fauna de vertebrados fósseis. O siltito tuffáceo do Membro Cerro Castaño, onde P. mayorum foi encontrado, foi depositado em ambiente fluvial em clima úmido com pouca sazonalidade e tem datação de de 101,62 milhões de anos [Krause et al, 2020].
(Crédito da imagem: Mariolanzas).
Acredita-se que o grande tamanho dos dinossauros fosse possível devido a uma combinação de algumas características, como [Hendry, 2023]:
- Pernas sob o corpo (e não abertas lateralmente) permitiam melhor sustentação e menor gasto de energia para se locomover. Além disto, os maiores dinossauros eram quadrúpedes.
- Pulsos e tornozelos menos flexíveis podiam ser mais fortes para suportar peso.
- Mãos e pés grandes e acolchoados ajudavam a distribuir o peso.
- Quadris largos aumentavam a estabilidade e proporcionavam mais espaço para o estômago.
- Pescoço longo facilitava alcançar a comida com menor esforço. Ele era possível por causa da cabeça pequena e do menor número de dentes, já que não era necessário mastigar para engolir.
- Ossos ocos e cheios de ar eram mais leves.
- Respiração eficiente por causa dos ossos cheios de ar associados ao sistema respiratório, em conjunto com os pulmões, aumentava a capacidade de absorver oxigênio continuamente e facilitava o crescimento do corpo.
- Metabolismo eficiente diminuía relativamente a necessidade de maior quantidade de comida. O grande estômago permitia que o alimento demorasse mais a passar por ele dando mais tempo para digerir e extrair nutrientes.
- O tamanho grande permitia:
- alcançar alimentos que estavam fora do alcance dos demais animais,
- sobreviver, na falta de alimentos, por mais tempo graças à gordura e à água armazenadas no corpo e
- viajar a mais longas distâncias e mais rapidamente em busca de novas fontes de alimento e água.
4.2.3.9. Extinção do Cretáceo-Paleógeno
No Cretáceo, o clima quente e predominantemente estável perdurou por quase 80 milhões de anos antes de terminar drasticamente, há 66 milhões de anos, com a extinção em massa do Cretáceo-Paleógeno [Scotese et al, 2024]. Esta extinção tem múltiplas causas seguidamente debatidas, entre elas regressão marinha,
vulcanismo massivo e impacto de asteroide [Archibald, 2002].
Uma das maiores regressões marinhas conhecidas é registrada no final do Cretáceo, há cerca de 65 milhões de anos, em curto período de dezenas ou centenas de milhares de anos. Estima-se que 18,2 milhões de km² de terra tenham sido expostos, o que corresponde aproximadamente à área da África atual. Grandes áreas de habitat costeiro foram alteradas e o tamanho das populações que ali viviam teria diminuído. Surgiram pontes terrestres permitindo migração de vertebrados terrestres com potencial aumento de competição entre espécies anteriormente separadas. Sistemas fluviais cresceram em extensão para alcançar as novas linhas costeiras, proporcionando maior habitat para organismos de água doce. O clima esfriou e os extremos climáticos aumentaram, estressando ainda mais o ambiente [Archibald, 2002].
A atividade vulcânica intensa ficou a cargo das erupções prolongadas das Traps de Deccan
que aconteceram na fronteira Cretáceo-Paleógeno com uma série de pulsos intermitentes curtos (de ∼100 mil anos) e com duas fases principais [Chiarenza et al, 2020]:
- uma que ocorreu na proximidade do final do Cretáceo (há ∼67,4 milhões de anos) e
- outra que começou pouco antes do limite Cretáceo-Paleógeno (há ∼66,1 milhões de anos) e continuou por ~710 mil anos no início do Paleógeno.
Foram liberados gases atmosféricos ativos, particularmente CO₂ e SO₂, produzindo um cenário que poderia levar a uma extinção em massa ou poderia estressar os ecossistemas globais tornando-os vulneráveis a agentes subsequentes. Incluindo neste cenário o impacto de um asteroide, as Traps de Deccan podem também ter atuado como agente amenizador dos efeitos catastróficos (decorrentes do impacto) sobre o clima e sobre os ecossistemas globais. Por exemplo, o vulcanismo de longo prazo, com o aquecimento induzido por dióxido de carbono, poderia ter reduzido a gravidade da extinção [Chiarenza et al, 2020].
Provavelmente, a causa mais importante da extinção no final do Cretáceo tenha sido um cenário de inverno extremo prolongado desencadeado pelo impacto do asteroide Chicxulub.

 Foto de 1969 do limite Cretáceo-Paleógeno na Baía de Loya, Hendaye, França, com rachadura desenvolvida exatamente no limite ao longo da camada de irídio.
Foto de 1969 do limite Cretáceo-Paleógeno na Baía de Loya, Hendaye, França, com rachadura desenvolvida exatamente no limite ao longo da camada de irídio.


Localização da cratera de impacto do Chicxulub na geografia atual.
(Imagem baseada em: GrandEscogriffe; NASA/JPL / David Fuchs / Mircalla22).
O impacto do Chicxulub teria feito as temperaturas despencarem devido à poeira e aos aerossóis que foram lançados na atmosfera absorvendo a luz solar [Scotese et al, 2024]. Assim, a vaporização do corpo celeste e do material impactado, juntamente com as cinzas de incêndios florestais, produziram um escurecimento atmosférico global que durou vários meses [Molina, 2013]. Houve resfriamento global com graves efeitos ecológicos em cascata [Chiarenza et al, 2020] provocando um colapso na cadeia alimentar e a extinção de ~75% de todas as espécies. O efeito deste evento nos ecossistemas globais foi superado apenas pela extinção do Permiano-Triássico [Scotese et al, 2024].
O registro paleobotânico mostra mudanças paleoambientais abrangentes, incluindo desmatamento e redução temporária brutal na diversidade, com algumas espécies oportunistas (principalmente samambaias) dominando durante o estágio de sobrevivência. Além disto, há evidências de chuva ácida e anoxia. Nas profundezas do oceano, houve acidificação que seria causa mais importante nas extinções do plâncton em calcário [Molina, 2013].
A fronteira Cretáceo-Paleógeno (também conhecida como fronteira K-T, referente a Cretáceo-Terciário) é identificada por meio de três critérios [Lyson et al, 2011]:
- diminuição de palinomorfos do Cretáceo sem recuperação subsequente,
- existência de um "pico de samambaias" e
- ocorrência de marcadores primários de impacto extraterrestre como a anomalia de irídio.
Um nível anormalmente rico em irídio coincidente com a súbita extinção em massa catastrófica teria sido produzido pelo impacto do Chicxulub. O estrato tipo desta evidência foi definido na seção El Kef, na Tunísia. Ela foi registrada em um intervalo de argila fina na fronteira Cretáceo-Paleógeno em diversos locais, como Gubbio na Itália, Stevns Klint na Dinamarca, Woodside Creek na Nova Zelândia, Caravaca na Espanha [Molina, 2013] e Baía de Loya em Hendaye na França [Mulder e Seibert, 2019]. Na Baía de Loya também foram observadas as últimas ocorrências e extinções de espécies de foraminíferos antes do limite Cretáceo-Paleógeno [Alegret et al, 2022].

Neste pequeno promontório, intercalada por margas do Cretáceo superior (abaixo) e calcário do Paleógeno (acima) [Mulder e Seibert, 2019], a camada de argila com irídio sofreu contínua erosão marinha que, em 2005, provocou previsível deslizamento de terra destruindo o afloramento [Thomas, 2016].
(Crédito da imagem: Jules Rhin).
Além da argila rica em irídio e da grande estrutura de cratera de impacto na Península de Yucatán, outras evidências permitem confirmar a validade da teoria do impacto, como [Molina, 2013]:
- microtectitos,
- espinélios ricos em Ni,
- quartzo chocado,
- níveis sedimentológicos produzidos por tsunamis,
- fluxos gravitacionais gigantescos e
- datação por 40Ar/39Ar em vidro de impacto.
A comprovação de que o impacto do Chicxulub e a extinção em massa do Cretáceo-Paleólogo foram contemporâneos é dada pela abundância e pelo pequeno tamanho dos foraminíferos planctônicos. Eles apresentam um padrão de extinção catastrófica e repentina que afetou mais de 70% das suas espécies e que se extinguiram repentinamente a partir do nível que continha as evidências do impacto [Molina, 2013].
Os ambientes mais afetados foram terrestres e marinhos pelágicos, onde a catástrofe foi tão repentina que não houve tempo suficiente para a seleção natural ocorrer, principalmente no chamado inverno de impacto.
O impacto produziu enormes tsunamis na área do Golfo do México, desestabilização e ruptura da plataforma marinha, gerando fluxos de massa de megaturbidito,
que em algumas seções cubanas se transformaram em enorme olistostroma,
atingindo dimensões de centenas de metros [Molina, 2013].
Das espécies marinhas, ~70% foram eliminadas. Para as plantas terrestres, os efeitos foram devastadores e, em especial, foi alterada a trajetória de evolução de diversificação das angiospermas, conforme ficou especialmente claro no desenvolvimento de florestas tropicais de estilo moderno [Benton e Sauquet, 2021]. Muitos grupos de animais que dominaram a Terra por 150 milhões de anos também foram extintos. Foram extintos os dinossauros não aviários, cujos potenciais habitats globais foram suprimidos [Chiarenza et al, 2020].
O debate sobre o dinossauro estratigraficamente mais jovem é antigo e envolve a lacuna de 3 m.
Acredita-se que o último dinossauro seja representado pelo fóssil de chifre de testa de um ceratopsiano
encontrado em depósito de planície de inundação de argilito siltoso, localizado a não mais de 13 cm abaixo do limite Cretáceo-Paleógeno, ou seja, dentro da lacuna de 3 m [Lyson et al, 2011].

Crânio do Triceratops prorsus
em exposição no Museu Peabody de Yale, em Connecticut, EUA.
O crânio deste ceratopsiano foi preservado na Formação Lance, em Wyoming, EUA [Farke, 2011] em cujos arenitos é encontrada abundante fauna vertebrada, assim como restos de plantas que indicam ambiente subtropical úmido. A fronteira Cretáceo-Paleógeno está contida mais próxima ao topo desta formação, mas é indefinida devido ao retrabalhamento dos sedimentos. Há elevação da concentração de irídio, mas não em intensidade elevada nem em estrato bem definido [Hope, 1996].
(Crédito da imagem: Longrich e Field).
Este evento de extinção em massa preparou o cenário para a efetiva evolução dos mamíferos [Scotese et al, 2024]. Esta catástrofe constitui uma das crises bióticas mais significativas da história da Terra e marca notavelmente a fronteira entre as eras Mesozoica e Cenozoica [Molina, 2013].
Nenhum comentário:
Postar um comentário